Glossary
| Aboral | Posterior | |
| Adult | (Sexually) mature | |
| Anal tubercle | Small tubercle above the last pair of spinnerets which closes the anus | 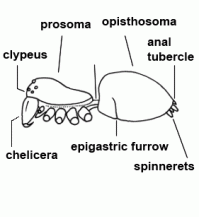 |
| Anatomy | The body of the spider is divided into two major divisions: an anterior part (prosoma) and a posterior part (opisthosoma) which are connected by a narrow stalk (pedicel). The extremities of the prosoma are (from anterior to posterior): chelicerae, pedipalps and 4 legs. The mouth opening is laterally bordered by the gnathocoxae, anteriorly by the normally soft skinned labrum and posteriorly by the sclerotised labium. The lateral areas of the prosoma are soft (thus not or only slightly sclerotised), whereas the dorsal cover (carapace, tergum) and the ventral plate (sternum) are sclerotised. The "head" part of the prosoma bears the eyes. The most important organs in the prosoma are 1 pair of poison glands, the brain (nervous system), esophagus and the sucking stomach, as well as the endosternite and the muscles. The opisthosoma bears on the anterior part of ventral side the epigastric furrow with the genital opening and the spiracles, posteriorly normally 3 pairs of spinnerets. Organs within the opisthosoma: the tube-shaped heart, ventrally the respiratory organs (book lung, tracheae) and the spinning glands. Furthermore are within the opisthosoma the testes or ovaries, respectively, the strongly segmented midgut and hindgut. The hairs on the body of the spider are sensory organs that response to mechanical or chemical stimulus | 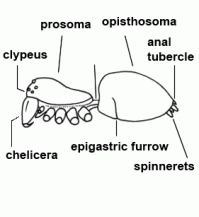 |
| Apodeme | A sclerotised infolding of the cuticula that provides an attachment for muscles. An important apodeme is situated under the fovea | |
| Apophysis | Excrescence, process. The various apophyses of the male palp play an important role during copulation. They can provide a secure coupling with the female's epigyne or vulva. They also act as an "internal" locking mechanism for the palp itself, so the individual parts of the bulbus can be fixed in an optimal position. | |
| Arachnids (Arachnida) | Class of 8-legged invertebrate animals; for example the orders Araneae (spiders), Scorpiones (scorpions) and mites (Acari) belong to this class. At present there are approx. 80.000 species known worldwide, however the effective number of species is probably much higher | |
| Artery | Vessel that carry blood away from the heart | |
| Arthropods (Arthropoda) | Phylum Arthropoda: e.g. the classes Insecta (insects), Crustacea (crustaceans), Myriapoda (millipedes) and Arachnida (arachnids) belong to this phylum. Over ¾ of all known species are arthropods. The >1 million species currently described are only a fraction of effectively existing species | |
| Atrium | Mostly relating to a vestibule of the vulva or the book lungs | |
| Autotomy | The voluntary separation of an extremity. Nearly all spiders can autotomise their legs. Autotomy is usually used when a leg of the spider is snatched by a predator or when an extremity cannot be freed during moulting. The site where a leg is autotomised lies usually between the coxa and the trochanter. A closing mechanism prevents leaking of the hemolymph after the leg has been detached. Autotomised legs can be replaced at the next moult, but they are considerably smaller than before. The normal leg size can be reached after further moultings | |
| Ballooning | In a certain stage of development and under certain weather conditions almost all spiders are able to "fly" or drift, respectively; for this they move to an elevated point, lift the opisthosoma and producing long strands of silk. These strands help them to take off even with a weak air current and quickly gain altitude (up to several 1000 m). For many spider species ballooning is the most effective way to colonise new habitats; they can cover a distance of several 100 km. Due to this aerial dispersal, spiders usually belong to the first settlers of new habitats | |
| Basal | Located at base | |
| Basal segment | First segment of the chelicera, mostly armed with teeth, bearing the fang | 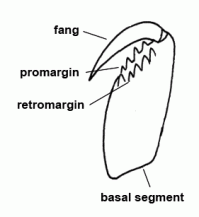 |
| Biotope preference | ||
| Book lung | Anterior respiration organ of most spiders located anteriolaterally of the epigastric furrow; the book lung consists of thin lamellae arranged like the leaves of a book. The opening is situated at the ventral side of opisthosoma. In dwarf-forms the book lung can be converted to tracheae | |
| Book lung trachea | ||
| Bristle | Hair-like formation of the cuticle. The term is usually used in determination keys for stronger and conspicuous, spine-like bristles on legs and palps | |
| Brood care | All female spiders exhibit brood care. In the simplest case, they protect the eggs, i.e., constructing a cocoon. The total amount of eggs inside a cocoon can vary from 100 up to 1000. The cocoon can be attached to a suitable place. Some spiders carry the cocoon until the spiderlings hatch (e.g. Lycosidae und Pisauridae, Scytodidae, Pholcidae) , thus they can provide optimal conditions for the development of the embryos. Many mothers tolerate the young spiders in their webs for some time, others (Lycosidae) carry the spiderlings on their body for some time. Only some spiders feed their spiderlings after hatching (e.g. Coelotes). In some Theridiidae species, the mother feed the spiderlings mouth-to-mouth with pre-digested, regurgitated food mash (regurgitation feeding), also known in Eresidae. In the latter case, the spiderlings feed on their dead mother | |
| Bulbus (genitalis) | Male copulatory organ on last segment of pedipalp; in entelegyne spiders, the bulbus consists of several flexible sclerites and membranous hematodocha in between; in haplogyne spiders, the bulbus is more simply developed, mostly uniformly globular or pear-shaped. See also cymbium | 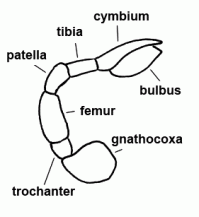 |
| Calamistrum | Organ that consists of one or two rows of bent bristles located dorsally on metatarsus IV. Only in cribellate spiders; used for combing out the silk from the cribellum |  |
| Carapace | Dorsal shield covering the surface of prosoma; named more accurately as "tergum" | |
| Cephalothorax | Anterior body part; see also prosoma | |
| Chelicerae | The most anterior pair of extremities of prosoma in arachnids, consisting of a basal segment and a fang | 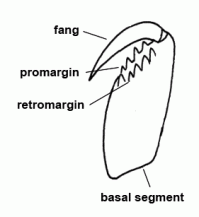 |
| Cheliceral furrow margins | Two distal margins of the basal cheliceral segment, mostly armed with teeth; the fang is situated between these two margins in resting state | |
| Chitin | Principal component of the cuticula; a nitrogenous polysaccharide | |
| Circulation | Spiders have an open circulatory system, i.e. that the hemolymph runs only a part of its way in blood vessels and is otherwise free floating among the organs. Spiders possess a rich system of arteries and only a few veins, though. The oxygen rich hemolymph flows in short lung veins from the book lungs to the dorsally situated tube-shaped heart. From there a large portion of the hemolymph is pumped into an aorta to the prosoma; a part of the hemolymph is also pumped into lateral and posterior arteries and reaches the opisthosoma. The flow of the oxygen-poor hemolymph back to the heart follows the gradient of decreasing pressure. Its course is not random but is guided along specific pathways | |
| Clavus | Backwardly directed, elongated process of median part of epigynal scapus; can be merged with parts of the epigyne, so that this "clavus sensu lato" bearing the copulatory openings | |
| Clypeus | 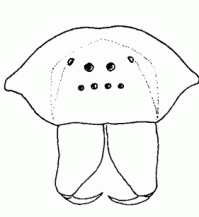 | |
| Cocoon | Egg-case; A wrapping for the eggs, consisting of special silk produced by the female. The shape of the cocoon is sometimes species-specific. The often thick and coloured silk layers protect the eggs to a certain amount against evaporation, fungal infection, egg predators and parasitoids). The finished cocoon is normally attached to a suitable place, sometimes also carried by the female (Lycosidae, Pisauridae, Scytodidae, Pholcidae). See also brood care | |
| Collecting and conserving of spiders | In scientific research, spiders are caught or collected with pitfall traps, yellow pans, Malaise traps, by beating and sweeping (with a net) the vegetation, sieving leaf litter and moss, etc. Small species can also be collected with an exhaustor, a spring steel forceps or with a moistened finger, larger species with a collecting container or by hand (in southern regions only recommendable with good knowledge of present species). Killing the collected spiders can be carried out with 70-80% alcohol, larger species can be anesthetised with vapour of ethyl ether and then put in alcohol. The containers with the collected spiders have to be labelled with date and locality to avoid later confusion. The medium for preserving the spiders is fresh alcohol (70-80%). The spiders can be stored in glass tubes closed with cotton wool, the tubes can be put in a larger, tightly closing jar filled with alcohol. Only specimens from one locality or from the same collecting date should be stored in a tube, or if possible, one species per tube only. If pedipalps and/or epigyne/vulva had to be detached (epigyne preparation), they should be put in a micro glass tube and stored together with the specimen in the same glass tube. Each tube should contain a label, written in Indian ink, or printed out with a laser printer on good quality paper. The label should contain: locality, grid reference, habitat, height above sea level, date, collector's name, maybe the name of the person who determined the species, slope of the locality, vegetation cover, etc. Nowadays, many databases are available, from which additional information can be gathered for the preserved specimens. Although, a database can never replace a good label, but rather complement. There are collections of spiders in many Museums of Natural History to familiarise oneself with the matter. If the collection is going to be published, voucher specimens have to be deposited in a museum to ensure a verification of the published data, so if there are doubts about the determination or taxonomic chances were made, anyone can easily check the voucher specimens. If someone abolish the reference collection of a study, no one can check the results, so one has to deal with being criticised for the unscientific work | |
| Colulus | Rudiment of the most anterior pair of spinnerets formed in embryonic development; its shape is mostly like a integumental lobe provided with bristles (not in all species visible or lacking completely) | |
| Conductor | Part of the embolic division, mostly serving as protection and guidance for the embolus | |
| Copulation | The male charges his bulbus with sperm before or during courtship. First, he spins a special sperm web and puts a drop of sperm on it. Then, he dips his palps into the sperm drop and the sperm is up taken. During copulation, the hematodochae (which are completely or partly reduced in haplogyne spiders) are inflated, so the sclerites of the bulbus are fixed in optimal position to each other and to the female genital organs. The sperm is transferred from the spermophor of bulbus through the embolus into the spermatheca of the female by increasing the hemolymph pressure and/or by displacement with glandular secretion. This procedure can last some seconds or up to some hours, depending on species. After the copulation, the female feed on the exhausted male (in some Araneidae, Theridiidae, etc.). Fertilization of the eggs takes place just before the eggs are deposited, often a long time after copulation | |
| Copulation | Mating | |
| Copulatory opening | External pore of the entrance duct in the epigyne of female spiders | |
| Courtship | Courtship can be defined as a certain behaviour of males (females as an exception) of a species which is signalling the willingness for mating to the other sex. Furthermore, this behaviour allows the females to recognise the male of the same species and the females can make some sort of evaluation of the quality of the male. Male spiders have often to avoid to be confused for prey. Thus, the courtship can also "calm" the female. The courtship behaviour of males is very various: many web building spiders court a female by vibrating the web of the female in a species-specific manner. Other spiders (e.g. many Theridiidae) produce acoustic signals with stridulatory organs, whilst others "dance" or showing waving movements. By doing so, they also display conspicuously coloured body parts. The males of the Lycosidae court with highly specific drumming patterns; the substrate can be caused to vibrate by pitching the legs, the palps and the opisthosoma onto the ground. The males of Pisaura court with a bridal gift: an insect wrapped in silk, on which the female feeds. Some morphological structures indicate chemical courtship signals. If the courtship succeed, the female takes up the mating position. Copulations aborted by females (e.g. in Pholcidae) indicate that during copulation "courtship signals", in a broader sense, also play an important role. Presumably, such signals provide information about the genetic quality of the mating partner | |
| Coxa | 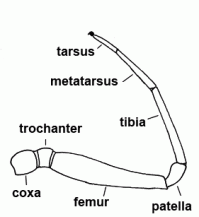 | |
| Cribellate catching threads | Catching threads produced by the cribellum. The very thin "catching wool" (diameter ca. 0.00001 mm) coats the more stronger axial threads. Prey is effectively entangled in this "wool", moreover, the cribellate silk has some adhesive properties | |
| Cribellate spiders | Spiders with a cribellum and a calamistrum | |
| Cribellum | A spinning plate derived from the first pair of spinnerets; only in cribellate spiders; this organ can produce up to 50.000 single threads. This "catching wool" is produced with the help of the calamistrum and is supported by a thicker axial thread in the web | 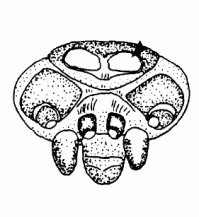 Cribellum |
| Ctenidia | Short process of the male palpal tibia of some Dictynidae species. The process is bearing very small and stout spines. | |
| Cuticle | The multi-layered stiff body surface of all arthropods; consisting of chitin and proteins; the outer layers of the cuticle are regularly renewed by moulting. Some internal organ (book lungs, fore- and hindgut) are also lined with cuticle | |
| Cuticula (Cuticle) | The external body shell of all arthropods, consisting of several layers. The main structural components are chitin and proteins. The cuticle is periodically renewed during growth (moulting) | |
| Cymbium | 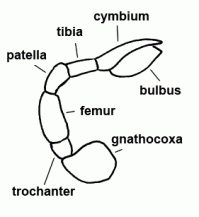 | |
| Digestion | ||
| Dissected epigyne | There is often the need for an epigynal dissection, in order to reliably identify a species. Therefore, a cut around the epigyne is needed. After this step, the epigyne with the inner vulva can be safely detached. Adherent tissue is carefully removed and the detached epigyne is put into clove oil or in an embedding medium which brightens the preparation (Svan, Berlese, Hoyer's). After a short time, the sclerotised structures of the vulva are visible and can be studied under the microscope. KOH or lactic acid cannot be recommended, normally the aforementioned procedure is sufficient. After examination, the detached epigyne have to be put into a micro glass tube and stored together with the specimen. The procedure is the same for haplogyne spiders, although more attention has to be paid, because an epigyne is missing and the vulva is often delicate and poorly visible from the outside | |
| Dissected vulva | Dissected epigyne | |
| Distal | Distant from centre or origin; see also proximal | |
| Distribution area | Area where a species occurs and reproduces. It may be compact (continuous) or not (disjunct). The area of a species is not constant, since species still may settle new areas within the postglacial colonization process. Additional dynamics are created by current anthropogenic processes such as modern climate change and landscape change. | |
| Distribution map | The distribution map of a species as given here per country or larger region bases primarily on data in Fauna Europaea (www.faunaeur.org). Additions are mentioned in the reference section. | |
| Dorsal | Faced opposite to the mouth (the back or upper surface); see also ventral | |
| Eggs | ||
| Embolic division | Distal part of bulbus, consisting of the embolus and mostly several additional sclerites | |
| Embolus | Sperm transferring organ of male bulbus; contains the opening of the spermophor which is situated inside the bulbus | |
| Endosternite | Inner skeletal element inside the prosoma of all spiders, formed by connective-tissue; serves as an attachment of several important muscles | |
| Enemies | Predatory insects (e.g. carabids, ants, kissing bugs) regularly prey on spiders. Although, other spiders are also important enemies, because many free hunting species feed on spiders ("cannibalism"). Pirate spiders (Mimetidae), for example, are specialised in preying on spiders. Among vertebrates toads, lizards and birds feed on spiders. Beside these predators, parasitoids are also important enemies. Also carnivorous plants, e.g. Drosera-Species (sundew) in bogs, catch wandering spiders | |
| Entelegyne spiders | ||
| Entrance duct | Connecting duct between copulatory opening and spermatheca of female vulva in entelegyne spiders; see also fertilisation duct | 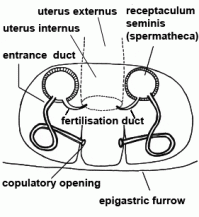 |
| Epigastric furrow | Transverse furrow on opisthosoma; entrance to the reproductive organs; the spiracles of the book lung are mostly situated lateral to the furrow | |
| Epigyne | Sclerotised external plate covering the female reproductive opening of female; with the copulatory opening for the embolus; only in entelegyne spiders | 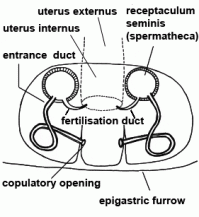 |
| Europe | Europe is biogeographically defined as western part of Eurasia. It does not include Canary Islands and other Atlantic islands close to Africa. The southern border of Europe is defined by the Mediterranean Sea. We include the Asian part of Turkey, Cyprus and the Caucasus states. In the east the border is indicated by Ural River and Ural Mountains. | 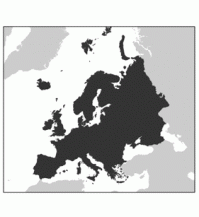 |
| Excretion | The extraintestinal digestion of the spiders and the subsequent filtering of the food mash are the reason why there is only a few amount of excretory products. The most important excretory organ is a pair of Malpighian tubules. These originate from the border between midgut and hindgut in the opisthosoma. The excrement is passed over the stercoral pocket to the anus. Excrement of spider mainly consists of guanine. In the prosoma of "primitive" spiders lie two pair of coxal glands, which are reduced to the anterior pair in most spiders. They open to the outside at the coxa of the walking legs and release a fluid only during feeding. Presumably, the coxal glands play an important role in ion and water balance. Furthermore, some gut cells and the nephrocytes in the hemolymph have excretory functions | |
| Exoskeleton | External rigid parts that serve as muscle attachments and support. The cuticula serves as exoskeleton in arthropods | |
| Extraintestinal digestion | Digestion occurring outside the intestinal tract; spiders sucking a pre-digested mash; digestive fluid from the intestinal tract is pouring on prey, so that its soft parts are dissolved (prey) | |
| Extremities | limbs; paired and originally segmented appendages. In spiders these are chelicerae, pedipalps and 4 pairs of legs; further modified extremities on opisthosoma (e.g. spinnerets) | |
| Eyefield | Part of head region bordered by the eyes | |
| Eyes | Normally, spiders have four pairs of eyes. According to their position, they are referred to as anterior and posterior median eyes (AME, PME) and as anterior and posterior lateral eyes (ALE, PLE). In some groups of spiders (e.g. Segestriidae, Dysderidae, Oonopidae, Scytodidae) the AME are reduced, so there are only six eyes. All eyes are ocelli (or so-called primitive eyes): beneath the cuticular lens lies a cellular vitreous body and the visual cells. A light-reflecting layer of crystalline deposits, the so-called tapetum, is characteristic for the ALE, PME and PLE of many spiders. Presumably, the tapetum allows seeing at night or in dim light. There is still a lack of knowledge about the performance of the visual sense in spiders. Generally, the eyes of the spiders are surely less efficient than those of vertebrates. It is known that Agelenidae and Lycosidae can perceive polarised light and use it for orientation. Some groups of spiders have especially complex eyes (e.g. the AME of free hunting jumping spiders); that indicates a highly developed visual sense (like e.g. the active "scanning" of objects, perception of UV light and a high resolution) | 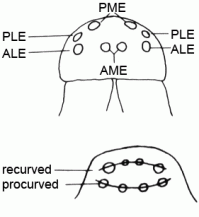 |
| Fang | Claw-like distal segment of the chelicera | |
| Fang | The claw-shaped movable terminal segment of the chelicerae; there is the opening of the poison gland at the tip of the fang | 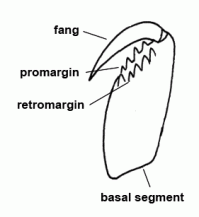 |
| Femur | Third segment of legs and palps | 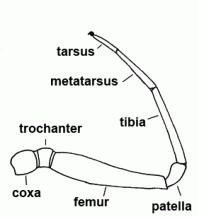 |
| Fertilisation duct | Duct with narrow lumen leading from spermatheca to uterus externus in the vulva of female entelegyne spiders; see also entrance duct | 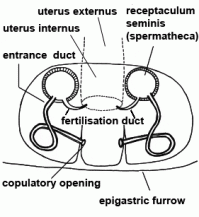 |
| Folium | Oak leaf shaped dorsal pattern on the opisthosoma; often with deposition of guanine, an excretion product (see excretion) | |
| Food | Spiders mainly feed on living arthropods, mostly insects. To a minor degree they can also feed on dead animals and eggs. The species of the genus Ero (Mimetidae) are highly specialised in preying on other spiders, Dysdera preferring woodlouses, Callilepis and Zodarion-species preferring ants. Presumably, pollen grains adhering to the sticky threads of orb webs play a role as a food source for spiderlings. The venomous bite paralyses the victim; the digestive juice is pumped into the wound by two pumps (pharynx and sucking stomach) and dissolves the inner organs of the victim. The liquefied food is filtered by specialised plumose bristles around the mouth and a so called palate plate. The palate plate possess fine chitinous platelets, which retains all particles > 1-3 micrometers and these particles are subsequently transported outwards. Therefore, only liquid food gets in the midgut, where it is stored and/or resorbed. Due to the extraintestinal digestion almost no solid particles can be found in the excrement. Many spiders chew their prey with the toothed cheliceral furrow margin, so it remains as a small clump. Some spiders suck out their prey through the fine holes caused by the fangs, so that an almost intact prey remains. 98% of the prey animal can be digested, only a small amount of an adequate prey remains (mainly cuticle of prey) | |
| Food uptake | Prey is held with the chelicerae and is intensely chewed. Only some specialists (e.g. Thomosidae) bite a small hole into the prey and suck out the victim through the bite hole. Simultaneously, the spider regurgitates digestive juice, which is produced in the midgut, so that the liquefied food can be sucked in. A system of small chitinous scales in the foregut, the so called palate plate retains all particles > 1-3 micrometers and these particles are subsequently transported outwards. Therefore, only liquid food gets in the midgut, where it is stored and/or resorbed. Due to the extraintestinal digestion almost no solid particles can be found in the excrement. Up to 98% of a prey can be eaten, the rest of the prey consists of chitinous parts (legs, etc.). The food mash reaches the numerous diverticles of the midgut where storage and resorption takes place. The volume of the opisthosoma can be considerably increased while intense food uptake | |
| Forehead margin | Anterior margin of the carapace | |
| Fovea | Dorsal, mostly longitudinal depression on carapace (often an attachment for muscles) | 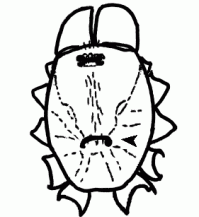 |
| Frontal | At the front | |
| Gnathocoxa | 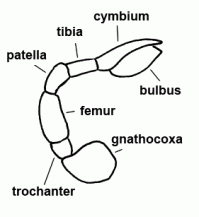 | |
| Guanine | Crystalline metabolic product of spiders; deposited in specialised cells under the cuticle and appears externally as white colour pattern (folium). See also excretion | |
| Habitat | Spiders are virtually found in all terrestrial habitats. Besides some unspecialised generalists (ubiquists), we know many species with highly specific preferences for some habitats. To what extend these specific preferences for some habitats in the nature match the actual requirements of a species is a current problem in research - presumably, some species are only displaced to extreme habitats by other species. These species aren't restricted to a certain habitat, in fact they only tolerate the conditions much better than other species. Beside competition, several other factors play a role in the choice of a habitat: the structural components, availability of prey, microclimatic conditions (temperature, humidity, shadowing, etc.). The effective habitat of a species is often realised at a small scale only. Important habitats in a forest are therefore the various soil layers, herb layer, as well as the shrub and tree layer (trunk, bark, treetops); even these narrowly defined areas aren't completely colonised by a species, but only where they find specific microclimatic conditions or other important circumstances. The current habitat of a species can rather be seen as a more or less dynamic arrangement of factors than a locally clearly defined area. This perspective can explain migrations of some species due to weather conditions or the occurring of a species in very different habitats depending on geographical position. There are examples of species which migrate from southern regions to Central Europe. They can occur in dense vegetation or even in forests in Mediterranean region, but in Central Europe they can only survive in very warm, open and vegetation-poor south-exposed slopes. Special cases are species which occur in buildings (synanthropic) or in ant nests (myrmecophilous). Only one European species, Argyroneta aquatica, can permanently live beneath the water surface | |
| Habitus | General appearance | |
| Haplogyne spiders | ||
| Hematodocha | Soft areas of the bulbus, inflatable when hemolymph pressure is increased; connects the sclerites of bulbus among each other, connects also cymbium and bulbus | |
| Hemocyanin | The bluish oxygen carrying pigment of the arthropods; binds the oxygen within the hemolymph (correspond to the haemoglobin of vertebrates) | |
| Hemolymph | Body fluid of the arthropods; from a functional point of view a mixture of blood and lymph fluid (name!) | |
| Interpulmonary furrow | Epigastric furrow | |
| Labidognatha | Spiders with chelicerae projecting downwards and fangs opposing each other; over 99% of native spider species are labidognathous. See also orthognathous spiders | |
| Labium | The lower lip, forming the posterior border of mouth opening | |
| Labrum | The upper lip, forming the posterior border of mouth opening | |
| Lamella (characteristica) | A lamellous sclerite on the bulbus of many sheetweb weavers | |
| Lateral | (situated) on the side | |
| Life cycle | The detailed life cycle of most spider species is unknown. Although, a rough pattern can be outlined after which many species, albeit not all, develop: the young spiders hatch ca. 2-3 weeks after the egg was laid. These young spiders aren't self-sufficient, they feed on yolk and possess in the first stage (formerly also named as prelarva) an incomplete leg segmentation. The second stage (larva) is already more differentiated, but leaving the cocoon not until a further moulting is accomplished into the first self-sufficient stage (nymph). The following 5-10 juvenile stages (frequently named as free stages) are mainly characterised by a continuous increase in size. In the last juvenile stage (subadultus), the sexual organs are formed under the cuticle, but only with the last moult the spiders do reach sexual maturity and have functional sexual organs. The males usually die soon after mating, though, the females of spiders which do live one year (life span) can survive several month, however. During this time they lay eggs at intervals of some weeks. The reproduction is normally restricted to a few weeks. The spiders overwinter, depending on species, as eggs, as juvenile stages or as adults. Therefore, there are several possibilities for seasonal niches, due to activity patterns (spring, summer or winter active spiders). The term activity time refers to the time in which adults occur, it coincides with the reproduction time | |
| Life span | Most spiders of temperate regions do live only one year; the half life span or more is allocated to the time before the final moult to an adult animal. In Nordic alpine regions, in which low temperatures slow down the life cycle, these species can also live two years. Larger spiders, e.g. many Gnaphosidae, Segestriidae, Eresidae or Araneidae in central Europe can live ca. 2-3 years. The females always live longer than the males. Orthognathous spiders (females) usually live much longer; Atypidae live up to eight years, tropical tarantulas even ca. 20 years, but there are also some small species which have several generations per year | |
| Lung | ||
| Median | In the middle or midline | |
| Median apophysis | 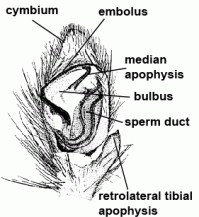 | |
| Metatarsus | Sixth segment of the leg, not found in the pedipalp | |
| Moulting | Casting off the old cuticle. Because of the rigid cuticle, spiders - like all other arthropods - have to moult from time to time, otherwise growth isn't possible. Normally, a spider passes through 5-10 moults to reach the adult stage. During the last moulting intervals the sexual dimorphism becomes apparent and after the final moult, the sexual organs are completely developed. Moultings after reaching the adult stage can only be seen in orthognathous as well as in few labidognathous spiders (Eresidae). First of all a new thin and strongly folded cuticle is formed under the old cuticle. During this process, the new cuticular sense organs (sensory hairs, slit sense organs, tarsal organs) are formed. A fluid provided with enzymes is secreted into the gap between new and old cuticle. The enzymes dissolve the inner parts of the old cuticle. The dissolved components are largely resorbed, so there is only little material lost in the moulting process. Due to an increase of the hemolymph pressure, the old cuticle is ruptured laterally at prosoma. The carapace is lifted off and the extremities are extracted afterwards, and then, finally, the opisthosoma. Also the organs with chitinous cuticle are moulted, like fore- and hindgut, the book lungs and the tracheae | |
| Nervous system | In spiders the nervous system is situated on the ventral side (unlike in vertebrates), thus ventral to the intestinal tract. In embryos there is still an original, segmental nerve cord. However, during the development all segmental ganglia and their transverse interconnections (commissures) migrate forwards into the prosoma, where they form a large and complex brain. The brain encloses the intestinal tract, therefore one can distinguish an (upper) supraesophageal ganglion (with the visual centre) and a (lower) subesophageal ganglion (with outgoing nerves of the extremities and the opisthosoma). Histologically distinct divisions of the central nervous system can be distinguished: a marginal layer of neurons (the cortex) and a central mass of nerve fibres (neuropil). Numerous neurosecretory cells are associated with the very efficient brain. The heart of the spiders possess an own ganglion | |
| Opisthosoma | Posterior body part, in spiders originally consisting of 12 segments (body structure) | 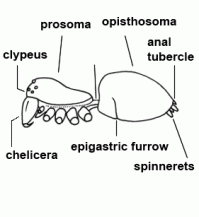 |
| Orthognathous | Presumably the ancestral position of the chelicerae; in orthognathous spiders the chelicerae are paraxial and mostly articulated vertically; typical for all mygalomorph spiders ("tarantulas"); most other spiders have chelicerae which are operating against each other (labidognathous), articulated horizontally like scissors | |
| Orthognathous spiders | Spiders with forwardly projecting basal segments of chelicerae and paraxially operating fangs. See also labidognathous spiders | |
| Palp | Second pair of extremities in Arachnida; usually divided into six segments in spiders. In females and juveniles leg-shaped, in adult males altered to a copulatory organ, often conspicuously coloured | 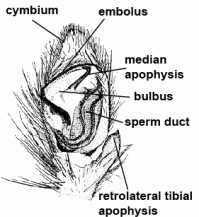 |
| Palpus | Palp | |
| Paracymbium | Structure that is completely or partly detached from cymbium | |
| Parasitoids | An organismn that has to spend a certain time of his life attached or within a (mostly larger) host organism in order to find optimal conditions for metabolism and/or reproduction at the expense of the host. Unlike parasites, parasitoids kill their hosts. Parasitoids of spiders are found among the hymenopterans (Hymenoptera). Pompilid wasps, but also some mud dobers (Sphecidae) paralyse spiders by their sting and the egg is laid on the spider's abdomen (the spider is still living). The hatched wasp larva is feeding on the living tissue until the spider succumbs. There are numerous egg parasitoids in the wasp family Ichneumonidae. They deposit their eggs in the spider's egg sac. Such egg parasitoids could already be understood true predators. Furthermore, there are also egg parasites among the flies (Diptera, e.g. the families Chloropidae, Sarcophagidae) and among the neuropteran (Neuroptera) the larvae of mantidfly (Mantispidae) | |
| Patella | "Knee"; fourth segment of legs and pedipalps | 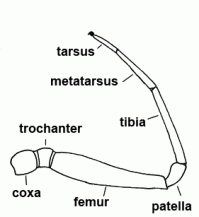 |
| Pedicel | Narrow stalk connecting prosoma and opisthosoma; from a developmental history point of view, it corresponds to the first opisthosomal segment | 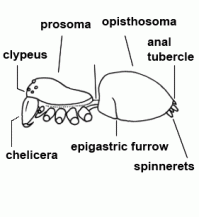 |
| Pedipalpus | ||
| Pharynx | The most anterior part of the digestive tract; there is a palate plate on the anterior wall of the pharynx, at which the food mash is filtered a second time (after a rough filtering by plumose bristles in the area of the mouth opening); levator and depressor muscles dilate or constrict the lumen of the pharynx, respectively, so that the pharynx operates as a pump for the food mash and the digestive juice. See also sucking stomach and food | |
| Phenology | Annual appearance of species, distinct developmental stages or physiological processes. For spiders, here the appearance of fertile (adult) individuals is give per month. | |
| Pheromones | Messenger substances which are produced in glands and secreted; they can be used for interspecific communication. Presumably, pheromones play an important role in finding the opposite sex in all spiders. They have only been detected for a few years. Adult females, which are ready for mating, secret pheromones on threads. These pheromones are presumably species-specific; there is only little known about their chemical structure | |
| Pigment | Dye | |
| Pleura | The lateral, mostly soft membranous area between carapace and sternum; extremities fan out of this area | |
| Preformed | To form beforehand | |
| Prey | Food | |
| Procursus | Retrolateral paracymbium found in pholcids | |
| Procurved | ||
| Prolateral | Situated on the front side of an extremity. See also retrolateral | |
| Prosoma | Anterior body part, consisting of six (merged) segments; extremities are attached to it | 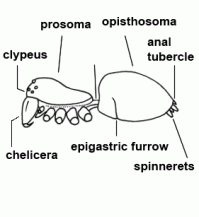 |
| Proximal | Close to the centre or origin; see also distal | |
| Pumping stomach | Sucking stomach | |
| Radix | A sclerite of the bulbus, situated basally to the embolus, pervaded by the spermophor | |
| Receptaculum seminis | ||
| Recurved | Eyes | |
| Regeneration | Autotomy | |
| Respiration | Most of the spiders possess one pair of book lungs consisting of stacks of lamellae like pages of a book. These lamellae are double-layered and filled with air, thus hollow. Small chitinous columns prevent collapse of the narrow air pockets. The distance between the lamellae, where the hemolymph circulates, is adhered by supporting cells. Oxygen from the air diffuses into the hemolymph and is bound to hemocyanin. In most spiders there is a pair of tracheae. Through this tubular respiratory organ the oxygen is directly reaching the organs. See also circulation | |
| Retinaculum | Mostly relating to a groove-like structure of the epigyne, sometimes also to single sclerites of the bulbus | |
| Retrolateral | Located at the back of an extremity. See also prolateral | |
| Scapus | Appendage of anterior epigynal margin | |
| Sclerite | Hardened area of the cuticle, due to deposition of sclerotin. See also scutellum/scutum | |
| Sclerotin | Tanned protein of yellow-brown colour, deposited into the cuticle (sclerotised cuticle), it provides toughness and resistibility | |
| Scopula | Tufts of hairs with adhesive function, located distally on the legs | |
| Scutellum/Scutum | A large sclerite on the opisthosoma | 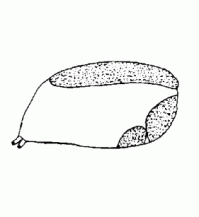 |
| Scuticula | See scutellum/scutum | |
| Segments | Successively arranged subdivisions of the body; in the original state characterised by a set of homogenous organs | |
| Sensory hairs | Most hairs and bristles are movable and articulated and have a mechanosensitive function. At their base there are sense cells which react to movements of the hairs. The extremely thin and usually very long trichobothria are only on legs and pedipalps and are extremely movable. They react to air currents and allows, as "remote tactile sense" some kind of "hearing". The short, steeply projecting, chemosensitive hairs on legs and pedipalps bearing a terminal pore opening, beneath which nerves end; they can be used, as contact chemoreceptors, for tasting and presumably for some kind of smelling | |
| Serrula | A row of densely adjoining tiny teeth on the gnathocoxa | |
| Sexual dimorphism | The genetically determined difference in form between male and female individuals of the same species. Beside the primary sexual characters (testes resp. ovaries with ducts leading to genital opening, not visible from outside), also the secondary sexual organs play an important role: Adult male spiders are easily recognisable by the thickened and modified tarsus of the pedipalp (bulbus), the females by the epigyne and/or vulva. Furthermore, adult males have almost always longer legs and in many Linyphiidae, the males have bizarre and often species specific head processes. Aside from a few exceptions, the males are smaller than the females and, during mating season, they vagabond more than females | |
| Sexual organs | ||
| Sigilla | Externally visible roundish oval muscle attachments | |
| Slit sense organs | Membranous slits in the cuticle with sensory function, they may occur singly or in groups ("lyriform organs"). A mechano-sensitive nerve is attached to the membrane. Compression of the slit elicites nerve impulses, so the measurement of the tension of the cuticle is possible. Slit sense organs respond to vibrations of the web, air or substrate, to movements of body parts relative to each other, to muscle activity, body weight and changes of the hemolymph pressure. Moreover, they play a role in kinesthetic orientation and interact central nervously with the trichobothria. Garden spiders possess over 4000 slit sense organs which are distributed over the whole body, but mainly on the legs. Small spiders possess about 250 slits sense organs | |
| Spermatheca | Mostly globular, club-shaped or barbell-shaped sperm storage organ of the vulva of female. See also copulation | 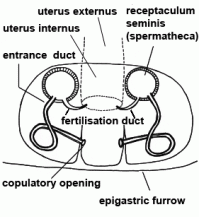 |
| Spermophor | Sperm duct, situated inside the bulbus, in which the sperm can be taken up, stored and expelled during copulation | 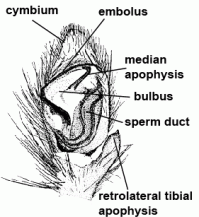 |
| Spider silk | All silk glands are located inside the abdomen and producing silk. Each gland leads to a specific spinneret, which opens to the outside in the form of a spigot. Spider silk consists of certain proteins (fibroins), which are enriched with pyrrolidin and salts (potassium hydrogen phosphate, potassium nitrate) to protect the silk from bacterial decomposition. The molecular ultrastructure of the silk threads is still unclear. Known so far, the proteins of the dragline consists to a great amount of the two amino acids glycine (ca. 42%) and alanine (ca. 25%). New models assume that the alanine-rich fractions of the thread occur in two forms: as crystals with a high degree of orientation and as protocrystals with a weak degree of orientation of the molecules. These both forms are embedded into a amorphous, glycine-rich matrix. More complex amino acids, like glutamine and others, affect the size of the crystals and the forming of chains, which connect the crystals with each other and with the surrounding amorphous matrix. It is assumed that the amorphous protein matrix accounts for the elasticity of the thread, whereas the crystalline fractions account for the tenacity of the thread. Old silk is eaten by e.g. the European garden spider. The proteins are nearly completely recycled by the spider's metabolism. After only 30 minutes 80-90% of the proteins of the eaten silk are again available for use | |
| Spinnerets | Phylogenetically, the spinnerets represent modified extremities of the 4. and 5. segment of the abdomen. Due to a division of the anlagen of the extremities into a smaller inner and a larger outer pair during embryonic development, 4 pairs of spinnerets emerge, which are only present in Mesothelae (see systematic). The anterior median pair is either reduced to a rudiment, the colulus, or to a cribellum. The spinning glands terminate in spigots on the surface of each spinneret. The Araneoidea (systematics) possess six different types of spinning glands and thus the greatest variety | 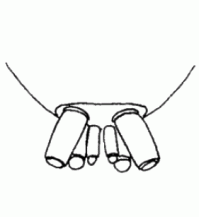 |
| Spiracle | The opening of the trachea on the ventral side of the body | 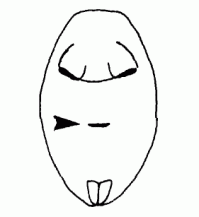 |
| Sternum | Ventral plate of the prosoma | 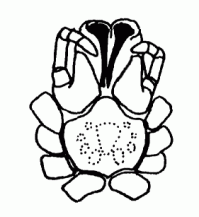 |
| Sticky threads | A catching thread provided with glue. Sticky threads are found in the orb webs of the Araneidae and Tetragnathidae, as well as in the webs of Theridiidae, Nesticidae, Linyphiidae, Theridiosomatidae, Mysmenidae and Anapidae. The glue originates from specialised spinning glands in the opisthosoma and coats a double silk thread as a thin layer. The glue condenses to fine droplets immediately after excretion | |
| Stridulation | Generating acoustical signal via a sharp edge which is rubbed against a ridge or area stuffed with humps, ridges or little cones. For generating sound, spiders rub e.g. humps on the pedipalp against the outer side of the chelicerae (many Linyphiidae), cones of coxa IV (Araneus) or a ridge at the posterior part of the carapace (Steatoda , Theridiidae) against the anterior margin of the opisthosoma | |
| Subtegulum | ||
| Sucking stomach | Specialised part of the foregut that can suck in pre-digested food mash or exhaust digestive juice; 2 pairs of muscles, attached to the endosternite and to the carapace, dilate the lumen of the sucking stomach resulting in a suction effect; circular muscles constrict the lumen and the content of the stomach is exhausted | |
| Synonym | Different scientific name that pertain to the same species or same phylogenetic group. According to the rule of zoological nomenclature, only the oldest published name for a taxon is valid (with a few exceptions). Here, the reference is always the newest version of The World Spider Catalog by N. I. Platnick | |
| Systematics | Worldwide there are now about 45.000 known species of spiders grouped into 114 families (WSC, 2014). The order can be divided into three suborders: (1) the Mesothelae (species-poor, only in Southeast Asia), (2) the Mygalomorphae ("tarantulas" s.l.) and the most species-rich (3) Araneomorphae. The Araneomorphae were originally cribellate, but the cribellum and the calamistrum were reduced in most forms. The phylogenetic relationships among the Araneomorphae are still unclear, but some groups can be characterised: presumably, the haplogyne spiders can be regarded as a monophyletic group. The Araneoidea comprise all spiders which can produce sticky threads (Araneidae, Tetragnathidae, Theridiidae, Nesticidae, Anapidae, Mysmenidae, Theridiosomatidae, Linyphiidae), as well as some groups which lost the ability to produce the sticky threads (Ero, fam. Mimetidae). Most likely, the orb web is an original construction from which all other webs within the Araneoidea are derived. Further, presumably "natural" groups are the Gnaphosids and their relatives (Gnaphosidae, Clubionidae, Corinnidae, Liocranidae, Anyphaenidae) and wolf spiders and their relatives (Lycosidae, Pisauridae, Oxyopidae). The cribellate spiders are no true "natural" group, but represent the "rest" of the Araneomorphae which keep the original Cribellum. Because the position of some families and genera within the Araneomorphae is very arguable and also nomenclature regularly changing, we decided to strictly follow the World Spider Catalog | |
| Tarsal organ | A sense organ situated on the tarsus; mostly, it forms a spherical pit on the dorsal surface of each tarsus; it mainly serves as a humidity and temperature receptor | |
| Tarsus | Seventh and last segment of the leg, sixth and last segment of the palp | 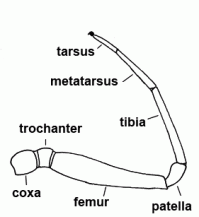 |
| Tegular apophysis | ||
| Tegulum | Largest median sclerite of bulbus; contains the spermophor | |
| Tergum | ||
| Terminal | At the tip, (situated) at the end | |
| Terminal apophysis | Apophysis located terminally on embolic division of bulbus | |
| Threats to spiders | Habitat loss and degradation are the most important threats to native spider species (in contrast to some tarantula species which are threatened by collecting). Partly, spiders are very closely associated to a certain type of microhabitat, which can be defined by various factors like climate, temperature, humidity, structural components and competition, etc. (see also habitat). Young spiders do often need different habitat types than adults, thus there have to be various adjoining macrohabitats (seasonal wandering). Generally, the closer a spider species is associated with a certain habitat, the more it is endangered. Only few, mostly disturbance-tolerating species had successfully colonised replacement habitats in the modern cultivated landscape. The emphasis on the endangerment is always linked to a decrease of the (total) population size. Rarity per se is only a potential measure of endangerment. Only in rare cases the decline of species can be documented, in most cases there is a lack of earlier data. Therefore, "Red Lists" resort to other criteria like the already mentioned association to a certain habitat and the endangerment of this habitats. The rarity (the decline, respectively) of a species can have various reasons: presumably, a rarely found species has very complex environmental requirements or only occurs in small, sparsely distributed or hard to reach habitats. Or microhabitats, which have been rarely studied due to methodical reasons (pitfall traps have been used for 40 years, studies of fallows have only been conducted for a short time). Without exact knowledge of the habitat preferences of such species, they will be only accidentally found and wrongly classified as "rare" | |
| Tibia | Fifth segment of the leg and palps | 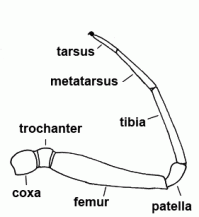 |
| Tibial apophysis | 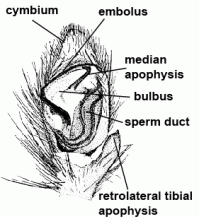 | |
| Trachea | Finely branching internal tubes, part of the respiratory system. Normally, the posterior respiratory organ of the opisthosoma in spiders, in some dwarf form also often the anterior one. See also respiration, book lung and spiracle | |
| Trichobothrium | A long, fine sensory hair on the legs and palps. The trichobothrium is embedded in a cup-shaped cuticular structure and reacts to air movements (also to acoustic pressure) with deflection whereby basal nerves are stimulated. Thus, some kind of "hearing" (perception of sound waves) is possible. The relative position of the trichobothria (e.g., on metatarsi of Linyphiidae) is often used as taxonomical character. | |
| Trochanter | The second segment of the leg or palps | 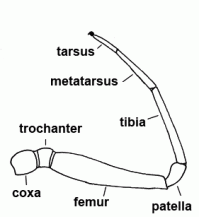 |
| Uterus externus | Outer part of uterus, in which the eggs are fertilised (in entelegyne spiders) | 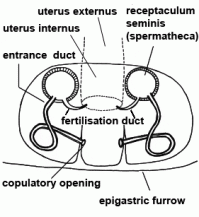 |
| Vein | A blood vessel that carries blood toward the heart | |
| Venom | All spiders, except for the family Uloboridae, possess a pair of poison glands located in the prosoma. Their adjoining ducts leading to an opening at the tip of the cheliceral fangs. When catching large and/or aggressive prey, the victims are bitten and venom is injected. Spider venom is a mixture of many components. It contains, amongst other compounds, effective neurotoxic polypeptides, thus insects are completely paralysed also after the injection of only a small amount of venom. The bite of Cheiracanthium punctorium (Clubionidae) can cause strong pain, fever and ague, however, effects of the bite aren't dangerous and a medical treatment isn't needed (except for allergic reactions which can also be caused by insect bites). Worldwide, only about 20-30 of the 40'000 spider species are dangerously poisonous to man | |
| Ventral | The side of the body where the mouth is located (belly or lower side). See also dorsal | |
| Vulva | Internal genital organ of female, consisting of the spermathecae and at least one duct between spermatheca and genital opening | |
| Webs | All spiders build webs for a more or less protected retreat; the orb weavers also use their web for catching prey. The very basal, recent spiders (the Mesothelae of south-east Asia) already build a silk-lined burrow and close the burrow with a very well camouflaged, silken lid. Other spiders use a silk-lined crevice under stones or bark or curled and silk-lined leaves as a retreat. There is an enormous variety in the shape of capture webs; their evolution is still not completely clear. Simple capture webs mostly consist of radial signal threads radiating from a retreat (e.g. Segestria). Agelenidae build a flat, slightly concave sheet web with a funnel-shaped retreat at one end (funnel webs). The cribellate threads and the sticky threads of the Araneoidea (e.g. Araneidae, Linyphiidae, Theridiidae) are specialised threads for catching prey. It is assumed that the possession of cribellum within the "higher" spiders (Araneomorphae) is the ancestral state; cribellum and calamistrum became reduced in many cases. Presumably, the Araneoidea also modified the orb web and replaced the cribellate catching thread with sticky thread. Later, the orb web itself was replaced with e.g. the various three-dimensional webs of Theridiidae and Linyphiidae. Other groups of spiders completely abandoned the web and now freely hunt their prey. See also systematics | |
| Webs | The spider web can serve as a residence or retreat; the webs often protect the spider with its cocoon from enemies; furthermore, many spiders use their webs for capturing prey. In the simplest case, a silk-lined burrow is used as a retreat with clean walls and balanced climate. Almost all spider species, as well as web spiders, build a silken retreat where they hide during the rest period. Such retreats can also be built under stones, under bark or in coiled up leaves. An expansion of the margin of the retreat leads to a funnel-shaped web (e.g. Agelenidae). The spider, which hides in the retreat, is alarmed when an insect enters the web; the spider comes out and subdues the prey. Furthermore, there are three-dimensional webs which are attached to the vegetation (e.g. in many Theridiidae). The horizontal sheet can be detached from ground and take up a central position (e.g. in many Linyphiidae). The horizontal sheet can also be reduced, thus a two-dimensional, highly symmetric web results (e.g. the orb webs of many Araneidae). There have been various modifications and reductions during the evolution, so there are many web forms and their phylogeny is still mostly unclear. The evolution of cribellate threads or sticky threads have to be happened several times independent of each other, presumably, just as the transition from three-dimensional webs to orb webs, as well as many web spiders in various families abandoned their webs and now capture their prey without webs |